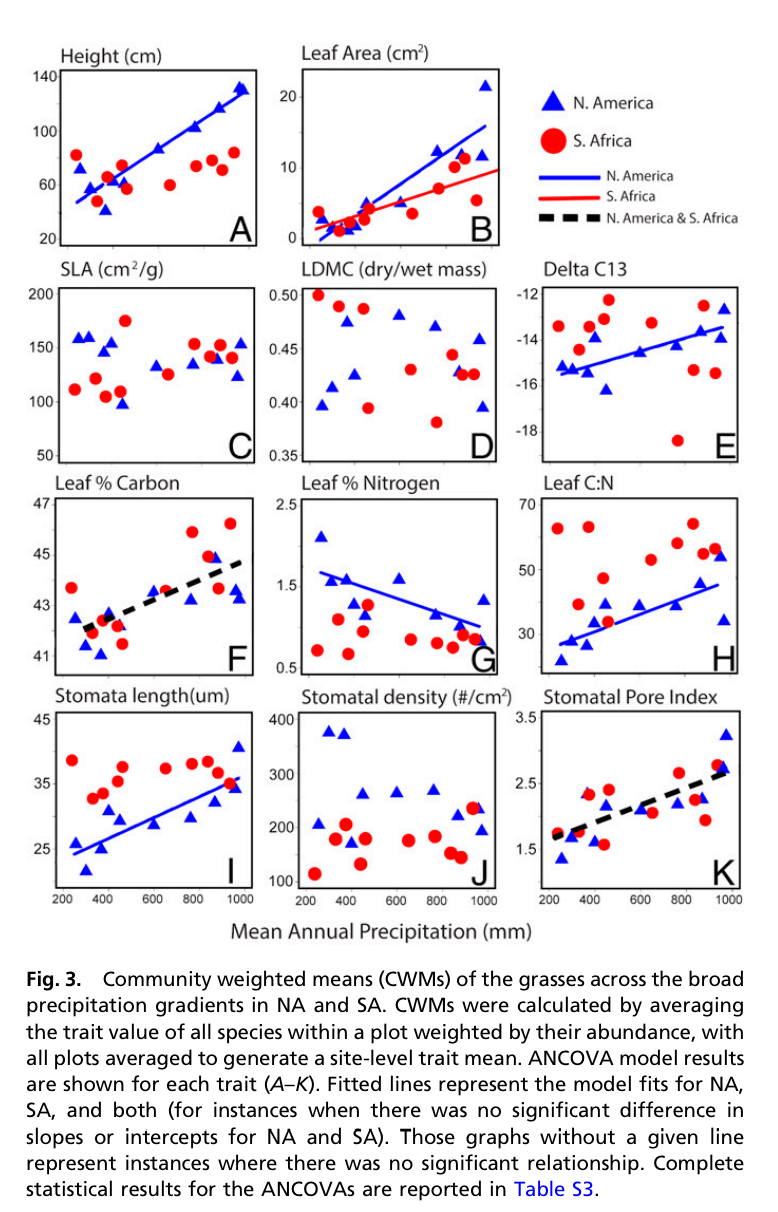
Contents of this post originally appeared on the Applied Ecologist, but with expanded thoughts here.
 |
Five years ago I spent my sabbatical in China and worked closely with a lab in Guangzhou. While there, I built meaningful collaborations and friendships that have continued to advance the science I'm involved with. While in China, I accompanied my friend, Jin-tian Li to a biodiversity field experiment on contaminated post-mining lands in Hunan province, and our discussions led to the just-published paper (please e-mail me if you want a copy) in the Journal of Applied Ecology, first-authored by a former PhD student in my lab, Pu Jia.
Why do we care about degraded lands?
 According
to the IPBES report on land degradation, the degradation of productive lands
and intact habitats is a major threat to sustainability, biodiversity and
ecosystem functioning globally, which reduces the resiliency of ecological and
economic systems. In many emerging economy countries, environmentally harmful
practices that result in contamination render lands and habitats seriously
degraded. In many circumstances, the restoration of contaminated habitats to
original conditions is not an option because the capacity for these habitats to
harbor intact native ecosystems is greatly compromised. In these cases, we need
management options that allow us to reclaim contaminated and degraded lands (Nathanail & Bardos 2005), and preferably
ones that increase biodiversity and ecosystem function (Rohr et al. 2016).
According
to the IPBES report on land degradation, the degradation of productive lands
and intact habitats is a major threat to sustainability, biodiversity and
ecosystem functioning globally, which reduces the resiliency of ecological and
economic systems. In many emerging economy countries, environmentally harmful
practices that result in contamination render lands and habitats seriously
degraded. In many circumstances, the restoration of contaminated habitats to
original conditions is not an option because the capacity for these habitats to
harbor intact native ecosystems is greatly compromised. In these cases, we need
management options that allow us to reclaim contaminated and degraded lands (Nathanail & Bardos 2005), and preferably
ones that increase biodiversity and ecosystem function (Rohr et al. 2016).
While the
ecological literature on the linkages between biodiversity and ecosystem
function is vast and rich (e.g., Tilman, Isbell
& Cowles 2014), the application of this field of research to
reclaiming contaminated lands has been strangely depauperate, and so there’s
little guidance on whether we should be planting diverse plant assemblages on
contaminated lands, or if we ought to simply plant the most productive species or those that provide
efficient phyto-removal of contaminants. This question is of fundamental importance to
places like China, where rapid development and industrialization through the
1970s-1990s resulted in severe contamination of lands near mining and mineral
processing facilities (Li et al. 2019), and now with China’s commitment to improving
it’s environmental health, biodiversity research has the ability to impact
policy and management at a national scale.
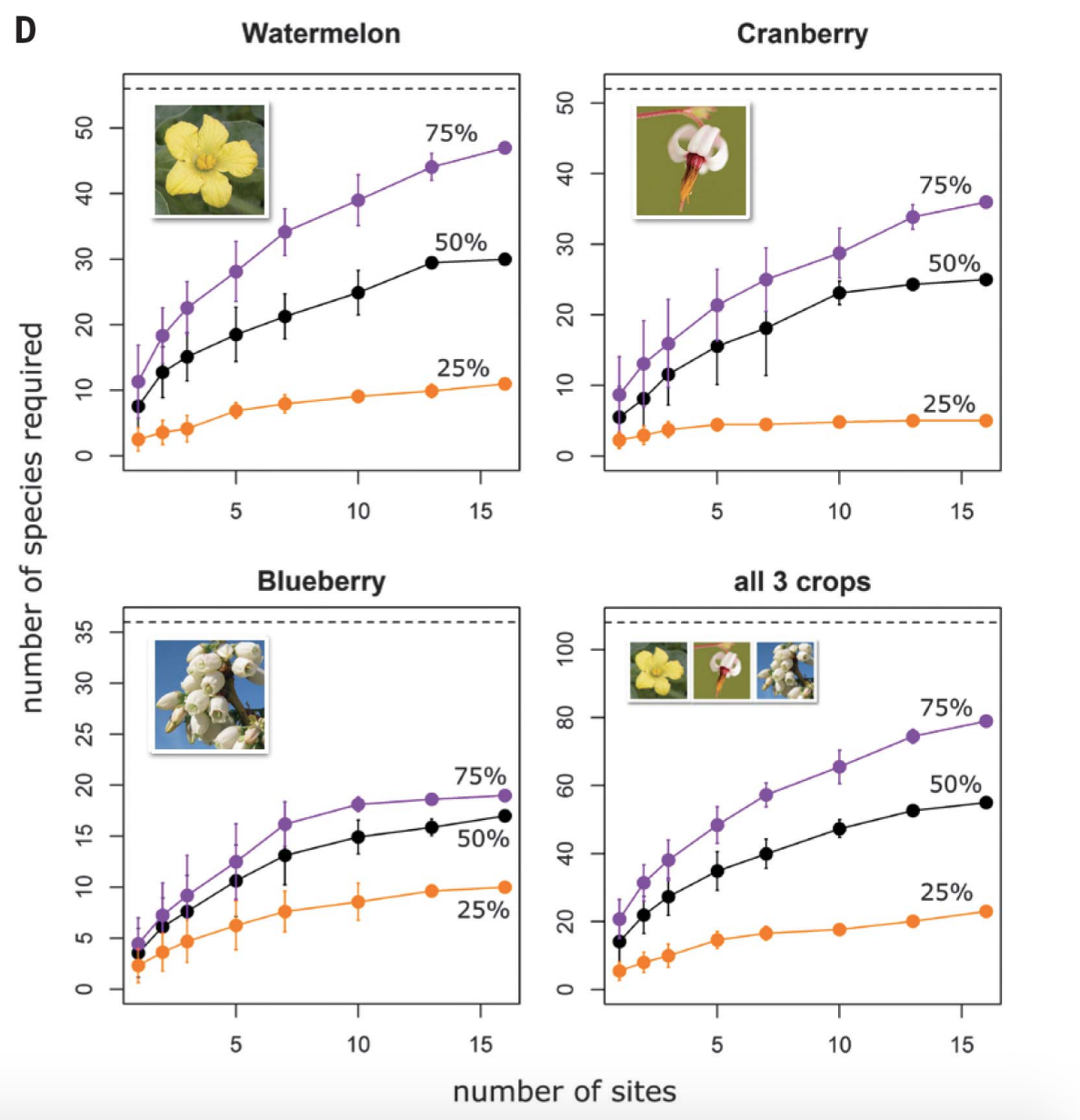
Our paper
We
evaluated whether more diverse plantings increased reclamation and ecosystem
functioning of a mine wasteland in Hunan Province, China, which had been
severely contaminated with cadmium and zinc over decades. We sowed plots with
1-16 species and these were selected from the herbaceous species that grew
around contaminated sites in the region, and more diverse assemblages produced
more biomass and were more stable over time. Further, there was less heavy
metal contamination of leaf tissues in the more diverse plantings, which
reduces the impact on herbivores.



Importantly though, plant diversity spurred plant-soil feedbacks (PSFs) that appeared to drive the increased ecosystem functioning. Higher plant diversity supported higher soil bacterial and fungal diversity. Importantly, higher plant diversity was accompanied with more soil cellulolytic bacteria that exude enzymes that degrade cellulose and so drive decomposition and nutrient cycling, which are essential components of a functioning ecosystem.
Furthermore, the multi-species assemblages also performed better because these high diversity treatments harboured fewer soil fungal pathogens (and by extension more beneficial soil fungi). This appeared to be driven by the fact that high plant diversity supported a greater diversity of soil chitinolytic bacteria that produce anti-fungal enzymes that degrade the chitin in cell walls of soil-borne plant-pathogenic fungi.
In the
search for efficient ways to reclaim contaminated lands, sowing high-diversity
plant assemblages appear to be an effective tool. The key for reclamation is
to ensure that soil processes like decomposition and nutrient cycling are able
to support a self-sustaining ecosystem, and higher plant diversity can ensure
this. The next steps will be to field test this in real reclamation projects and to see this research work its way into best practices.
Li, T., Liu, Y., Lin, S., Liu, Y. &
Xie, Y. (2019) Soil pollution management
in China: a brief introduction. Sustainability, 11, 556.
Nathanail, C.P. & Bardos, R.P. (2005) Reclamation of contaminated land. John
Wiley & Sons.
Rohr, J.R., Farag, A.M., Cadotte, M.W.,
Clements, W.H., Smith, J.R., Ulrich, C.P. & Woods, R. (2016) Transforming
ecosystems: when, where,
and how to restore contaminated sites. Integrated
Environmental Assessment and Management, 12, 273-283.
Tilman, D., Isbell, F. & Cowles, J.M.
(2014) Biodiversity and ecosystem
functioning. Annual Review of Ecology,
Evolution and Systematics, 45,
471-493.